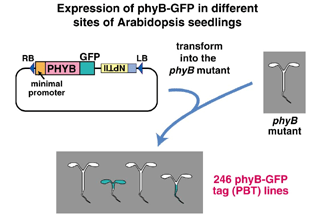
フィトクロムやクリプトクロムなどの光受容体はほぼ全身で発現している。器官毎の光受容を調べるには、部分照射実験や、器官切除実験が有効であるが、組織毎の役割をこのような方法で調べるのは困難である。そこで我々は、シロイヌナズナのphyB欠損変異体にphyB-GFP融合タンパク質遺伝子を導入し、エンハンサートラップ法の原理に基づいて、phyB-GFP融合タンパク質を組織特異的に発現する系統を多数、作出した(Endo et al., 2005)。
 * 葉肉phyBによる花芽形成制御
* 葉肉phyBによる花芽形成制御
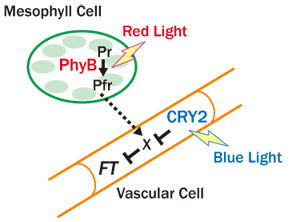
シロイヌナズナではphyBが活性化されると花芽形成が遅延する。この結果、いわゆる避陰反応のひとつとして、他の植物の陰では花芽形成が早まる。上記の系統を用いて、どの組織のphyBが花成を遅延するか調べたところ、葉肉のphyBにのみ、そのような活性があることが分かった。ここで、phyBによる花芽形成遅延においては、花芽形成の鍵因子であるFT遺伝子の発現が抑制されることが知られていた。そこで、上記の系統でFT遺伝子の発現を組織を分離して調べたところ、葉肉のphyBは維管束におけるFT遺伝子発現を抑制していることが分かった(Endo et al., 2005)。以上の結果は、葉肉細胞から維管束細胞への未知のシグナル伝達が存在するこを示している。
* 維管束cry2による花芽形成制御
シロイヌナズナではphyBに対してcry2が拮抗的に働き、花芽形成を促進することが知られている。また、cry2の下流でもFT遺伝子の発現が変化する。そこで、cry2-GFPを様々な組織特異的プロモーターを用いてシロイヌナズナのcry2変異体で発現させ、もともとの花成遅延表現型が相補されるかどうか調べた。その結果、phyBの場合とは対照的に、cry2-GFPは維管束で発現させたときのみ効果的であった。また、予想通り、維管束のcry2-GFPの働きで維管束のFT遺伝子の発現が促進された(Endo et al., 2007)。以上の結果は、同じ光受容体でありながら、phyBとcyr2は別の組織で花芽形成を制御していることを示している。
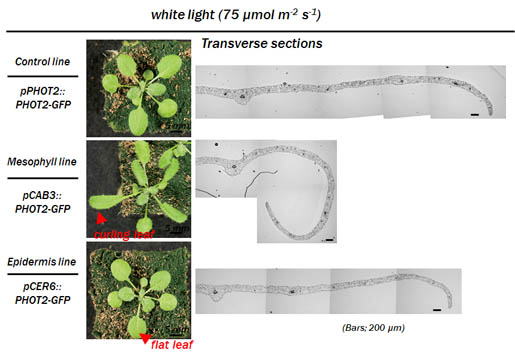
 * 葉肉細胞のフォトトロピンによる細胞自律的な柵状組織形成
* 葉肉細胞のフォトトロピンによる細胞自律的な柵状組織形成
我々はフォトトロピンが、葉の扁平化に加えて青色光による柵状組織分化を促進することを見出した。さらに、フォトトロピンを表皮または葉肉組織で発現させてその応答を観察したところ、柵状組織の分化は、葉肉細胞のフォトトロピンの細胞自律的な働きにより制御されていることが分かった(Kozuka et al., 2011)。
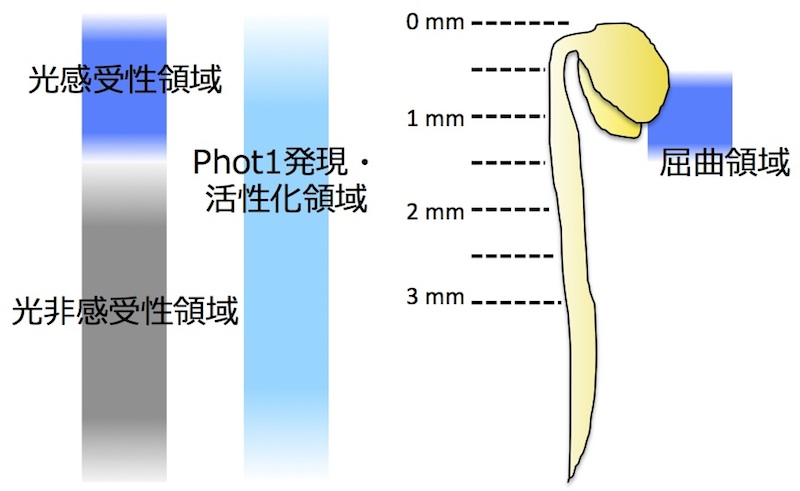
 * 光屈性における空間的制御機構
* 光屈性における空間的制御機構
シロイヌナズナを材料に、双子葉植物の光屈性が、「どこで光を感知し」、「どこで情報伝達が担われ」、「どこで屈曲が起こるか」を詳細に調べた。その結果、シロイヌナズナでは光感受性部位と屈曲部位の両領域が芽生えの上部に限定されることを見出した。従って、幼葉鞘の先端部で光を感知し基部が屈曲するイネ科植物芽生えと異なり、双子葉植物の光屈性はより部位自律性が高いことが明らかとなった(Yamamoto et al., 2013)。
 個体の光応答 研究topへ
個体の光応答 研究topへ